The sensory and motor roles of auditory hair cells Nature Reviews Neuroscience
Table Of Content
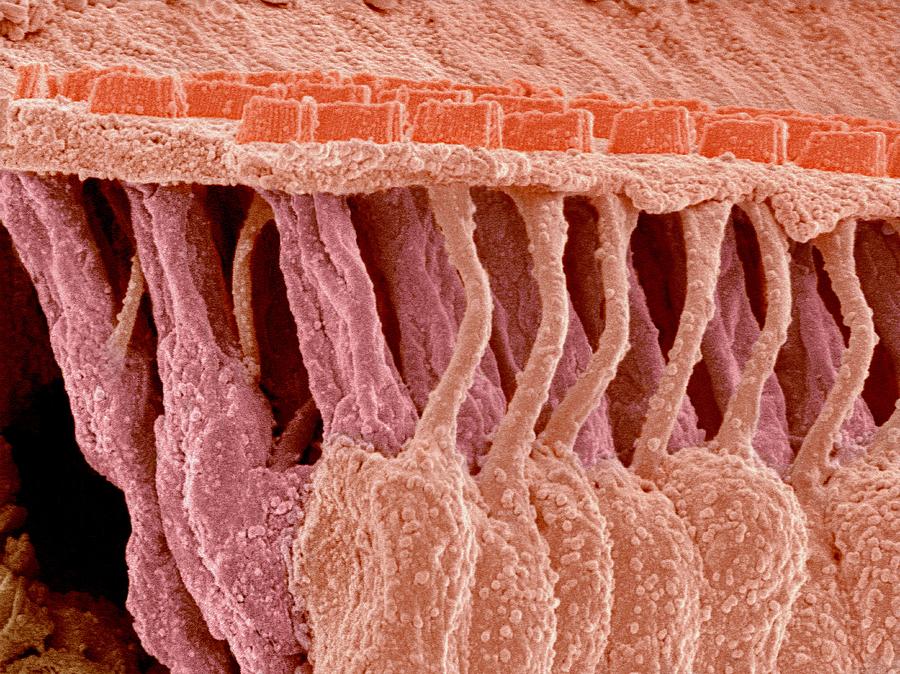
In the organ of Corti, HCs are categorized as inner hair cells (IHCs) and outer hair cells (OHCs) (Figure 1; Atkinson et al., 2015). Every HC is supported by several highly specialized cells, such as Deiters’ cells, pillar cells, inner border cells, and Hensen’s cells. All of the HCs are highly differentiated and polarized, and each acts as a mechano-electrical transducer that turns physical signals into electrical signals. External sensory stimuli physically open MET channels leading to an influx of K+ ions that depolarizes the HC. HC depolarization activates Ca2+ channels at the plasma membrane resulting in neurotransmission onto spiral ganglion neurons via the cochlear nerve. Finally, physical signals turn into electrical signals, which then pass through spiral ganglion neurons via the cochlear nerve, and the sensory signal ultimately reaches the cortex via the auditory pathway (Fettiplace, 2017; Ashmore, 2020).
What common conditions and disorders affect the inner ear?
Both type I and type II hair cells form ribbon synapses onto the primary afferent, and it is notable that one primary afferent may form synapses with both type I and type II hair cells (Eatock and Lysakowski 2006). However, the presence of a calyx constrains the outward diffusion of K+ from the basolateral membrane of the type I cell, leading to accumulation of K+ in the synaptic cleft. Consistent with this idea, the calyceal afferent membrane contains voltage-sensitive KCNQ-type channels, providing both a Nernstian permeability pathway and an exit channel from the synaptic cleft for accumulated K+ (Songer and Eatock 2013). The diagnostic feature of sensory hair cells is the presence of an apical bundle of stereocilia, known as the hair bundle.
Chapter 36: Plant Responses to the Environment
It is sometimes used with other ingredients like coal tar and zinc pyrithione to soften dry, scaly skin. Exfoliating products, which remove dead skin cells from the top layer of skin, can help keep the scalp healthy and clean. These products might also prevent hair follicles from clogging and remove hair products, oil, debris, and dead skin buildup. Finally, if you’re applying hair growth serum in the evening, consider your skin type—since certain ingredients, such as oils, can potentially rub off onto your pillow and be transferred to skin. “For sensitive or acne-prone skin, either frequent pillowcase changes or wearing a hair bonnet after serum application may be helpful,” she says.
Synaptic transmission
Collagen, which makes up bones and connective tissue, is another protein that resists decay. They both have amino acid compositions that lend to their strength (although they have several differences in their amino acid composition). Both are linear and tightly coiled proteins with multiple polypeptide subunits. Hair outlasts most other soft tissues due to the insoluble and stable structure of keratin. Its chemical composition makes it difficult for proteolytic enzymes – enzymes that break down proteins – to break down hair.
rs.onload = function()
When the cilia move toward the shortest cilium, the tip lengths are compressed and the cation channels close hyperpolarizing the cell, making it more negative inside, decreasing neurotransmitter release. If you have sensitive skin, consider using a mild product with natural ingredients or one designed for sensitive skin. When someone brushes a hand across your skin, it’s like a breeze blowing through a forest of countless small hairs. Nerves that surround your hair follicles detect that contact, and very far away in your brain, other cells fire. Some of the neurons responding to light contact might make you shiver and give you goose bumps. Development of hair cell phenotype and calyx nerve terminals in the neonatal mouse utricle.
How to Detox Your Scalp Safely at Home

Two other proteins, myosin XVa and whirlin, are also located at stereocilia tips, and all three are required for normal stereocilia length regulation. Myosin XVa functions in part by transporting EPS8 to the stereocilia tips (Belyantseva et al. 2005; Manor et al. 2011). Diagram of a stereotypical vertebrate hair cell, showing the major features of the cell and its relationship to the supporting cells around it.
Under fortunate conditions, this leaves enough intact tip links to allow electrical recording from one or a few channels at a time, and thus measurement of the channel conductance. When the extracellular Ca2+ concentration was subsequently raised to about 3 mM, the channel conductance measured was on the order of 100 picosiemens (100 pS) (Ohmori 1985; Crawford et al. 1991; Beurg et al. 2006). In contrast, most voltage-gated ion channels have conductances in the range of 10–30 pS (Hille 2001). Even more surprising, the conductance of the mechano-sensitive channel nearly doubled when the extracellular Ca2+ was lowered to its normal (sub-millimolar) endolymph concentration (Crawford et al. 1991). The increased conductance in low Ca2+ suggests that Ca2+ has an inhibitory effect on the channel (Beurg et al. 2010).
Myosin VI has also been implicated in adaptation (Marcotti et al. 2016). The appropriate mechanical stimulus for a hair cell is one that displaces the stereocilia bundle in the direction of the tallest stereocilia. This was demonstrated by Hudspeth and Corey using an isolated bullfrog saccular epithelium preparation (Hudspeth and Corey 1977). Minute displacements of the hair bundle produced depolarizing receptor potentials of several millivolts, and displacement in the opposite direction caused a hyperpolarization, though it was smaller in amplitude. Similar experiments on mammalian outer hair cells indicate that a 250 nm deflection of the hair bundle generates a half-maximal activation of the inward current and a 500 nm deflection is saturating (Fettiplace and Kim 2014).
Itchy Scalp
Top 3 grants in regenerative medicine: January 2024 - RegMedNet
Top 3 grants in regenerative medicine: January 2024.
Posted: Tue, 06 Feb 2024 08:00:00 GMT [source]
The 'hair' bundle on the apical surface of these cells is a mechanosensitive organelle that consists of precisely organized actin-filled projections known as stereocilia. Alterations in hair-bundle morphogenesis can result in hearing loss, balance defects or both. Auditory hair cells also have structural features that vary tonotopically along the length of the basilar membrane in the mammalian cochlea. Hair cells toward the apex, which are most sensitive to low frequencies, have longer stereocilia, and hair cells toward the basal end, sensitive to high frequencies, have shorter stereocilia.
Once tension reaches the point for channel opening, Ca2 influx would again act to reduce the tension. In this way, a negative feedback loop would keep the channels optimally positioned for sensitivity (Hudspeth et al. 2000; Farris et al. 2006). Recent evidence in support of this model is that directed mutation of the myosin1c gene, combined with an ADP analog that interferes with the mutated myosin, selectively blocks slow adaptation in the transgenic mice carrying the mutation (Holt et al. 2002). It has also been reported that myosin VIIa is necessary for slow adaptation in mouse cochlear hair cells (Kros et al. 2002).
These differences affect the passive compliance of the stereocilia, to which the opening and closing of the mechanosensitivie channels contribute an active component (i.e., greater compliance when the channels open and reduced compliance when the channels close). The features contribute mechanical tuning to hair cells that adds to their electrical tuning (Fettiplace and Ricci 2006). Mature hair cells do not produce action potentials, so all synaptic transmission is based on graded receptor potentials. Hair bundle displacement produces inward currents as large as 10 pA for 1 nm of displacement, which would lead to a depolarization of 1 mV for a hair cell having an input resistance of 100 MΩ (Fettiplace and Ricci 2006). The physiological range for receptor potentials is from the resting potential to about −20 mV (Glowatzki et al. 2008), corresponding to hair bundle displacements of up to 50 nm under physiological conditions.
In most reptiles, the ear develops further with an internal eardrum, giving rise to a prototypic external auditory canal (Schwab et al., 2020). While the ears of birds and mammals differ greatly, they still have highly developed outer, middle, and inner ears. The cochlea of mammals provides a single organ responsible for hearing that can cooperate with the other sensory functions of the ear. The longest stereocilia and related structures have a very close interrelationship, so the complete degradation of cochlear HC kinocilia will not have a profound physiological effect. The kinocilium shows a 9 + 2 axoneme pattern with typical motile cilium structures such as outer dynein arms and radial spokes. However, a lack of inner dynein arms renders the lack of motor function.
According to the World Health Organization (WHO), hearing loss affects more than 400 million people around the world today. Humans are unable to reverse the effects of hearing loss, but a biological process found in other animal species may hold the key to reversing this widespread problem. Zoe F. Mann and Matthew W. Kelley at NIH’s National Institute on Deafness and Other Communication Disorders (NIDCD) suspected that a molecular concentration gradient may guide the cells during development.
Like numbers on a ruler, the cell positions might be marked by levels of a signaling molecule. When the cilia move towards the tallest cilium, the tip lengths stretch and pull open attached cat ion channels. Potassium ions flow into the cell, depolarizing it, making it more positively charged inside. The best way to prevent scalp buildup is to stop it before it even starts.


Comments
Post a Comment